
安徽师范大学附属中学关于学生科技论文格式的规定
附件下载
为保证论文的质量,统一论文的撰写、印刷,便于处理、储存、检索、交流、传播,特制定本规定:
一、论文格式:
1、“标题”方正小标宋_GBK小二
2、“作者”楷体小四居中
3、“摘要”加方框,黑体五号
4、摘要内容楷体五号
5、“关键词”加方框,黑体五号
6、关键词内容楷体五号
7、正文内容宋体五号;行间距统一为:最小值20磅
8、标题层级的编码方法为:
l 文史类编码按照“一、二、三、四、……”序号字体格式
第一级用一、二、三……依次编码,一级标题后加顿号(、)与标题空开;黑体四号
第二级用(一)(二)(三)……依次编码,二级标题后不加点号;楷体五号
第三级用1. 2. 3.……三级标题后用点(.)与标题分开。仿宋五号
l 理工、体育类用阿拉伯数字编码,层次格式为:
1××××(黑体四号,居左)××××××××××××××(内容用5号宋体)
1.1××××(楷体五号,居左)×××××××××××××(内容用5号宋体)
1.1.1××××(仿宋五号,居左)××××××××××××××××××××(内容用5号宋体)
二、参考文献:
1、“参考文献”加方框,黑体五号居中
2、参考文献内容
(一)参考文献以文献在整个论文中出现的次序用[1]、[2]、[3]……形式统一排序、依次列出。
(二)参考文献的格式:
著作:[序号]作者.译者.书名.版本.出版地.出版社.出版时间.引用部分起止页
期刊:[序号]作者.译者.文章题目.期刊名.年份.卷号(期数).引用部分起止页
会议论文集:[序号]作者.译者.文章名.文集名.会址.开会年.出版地.出版者.出版时间.引用部分起止页
三、范例:
不同地理居群冰清绢蝶翅脉的几何形态学初步分析
(1.安徽师范大学附属中学,安徽芜湖241000;2.安徽师范大学附属外国语学校,安徽芜湖,241000. 3.安徽师范大学生命科学学院,安徽芜湖241000)
[摘要]本文对我国冰清绢蝶主要地理分布区的10个地理居群200个体翅脉脉相的相关指标进行测量统计和聚类分析,以期探讨几何形态学方法在研究蝶类昆虫种内居群间分化的可行性和有效性。利用三维数码显微镜测量分析了前后翅共计17种脉相的几何形态学性状,应用SPSS 21.0软件对筛选出的有效性状进行了统计和聚类分析显示:冰清绢蝶10个地理居群的形态分化和它们的地理分布之间存在明显的相关性;安徽滁州琅琊山的居群在聚类树上没能显示出与其地理分布间的明显关联性。研究表明,几何形态学分析方法在蝶类的种内分化研究中是一种十分有效的辅助手段。
[关键词]冰清绢蝶;地理居群;翅脉特征;几何形态学;系统分类
在传统昆虫分类学中,昆虫的翅脉特征是一个非常重要的分类学指标。它不仅对昆虫的分类鉴定有重要意义,还可以应用于对昆虫的物种多样性、系统发生发育及昆虫在不良环境条件下所发生的变异程度的研究[1]。
几何形态学(Geometric morphometrics)是关于形态定量比较的科学计算工具,是用数学方法描述或分析一个图像形状结构的方法。其研究方法多种多样,但在技术路线上都需要先将形态数据数字化,再比对不同的样本,通过比对计算出样本之间的形状和大小差距,意在找到样本之间的“同源信号”和“简约信息位点”。通过比对筛选出有效生物学性状,再用统计学的方法统计聚类。在国外,几何形态学方法已广泛应用于探讨生物的个体发育、种群分化、系统进化等多个方面,研究对象以大动物居多。在国内,仅用于昆虫的分类鉴定及远程自动识别研究中。目前国内随着计算机图像处理技术的不断发展,以昆虫翅脉特征提取为核心技术的软件也日趋完善,图像数字化处理技术与传统昆虫形态学结合已经充分应用到昆虫种间的分类鉴定中[1-5],但是运用几何形态学方法研究蝶类昆虫种内居群间分化国内外鲜有报道。
绢蝶属(Parnassius),是绢蝶亚科绢蝶族中的一个属,物种繁多,广泛分布于古北界及新北界温带的高山地区,从巴基斯坦到中亚和中国,部分物种扩至到欧州、日本及北美洲,因物种分布广,各地理因素如山脉、海洋、河流等导致物种隔离,继而演化出许多亚种[6]。我国绢蝶资源丰富,据周尧《中国蝶类志》记载:全世界绢蝶有38种,仅我国就有35种,124亚种。主要产于高山上,耐寒力强,少数种类分布在低海拔,寄主植物是景天科及紫堇、延胡索等。冰清绢蝶(Parnassius glacialis Butler)是分布在低海拔地区的绢蝶,也是我国江南分布的唯一一种绢蝶,这可能是第四纪冰期的遗留物种[7],分布于河南、山东、陕西、湖北、安徽、浙江、江苏、甘肃等地。寄主植物单一,主食罂粟科紫堇属的延胡索(Corydalis yanhusuo )和小药八旦子(Corydalis caudata)[6,8,9]。其形态学特征:体黑色,披有黄色长毛,颈部有1轮黄色毛丛。翅色白而稍呈蜡黄,翅质薄而半透明,温润如丝绢,翅脉灰黑褐色。前翅亚外缘有1条褐色带纹,中室内和中室端各有1个黑褐色斑纹,后翅内缘的黑带较宽。(图1)[7,10]。
生物地理学是生物学和地理学间的边缘学科,研究生物在时空上的分布及阐释物种的地理分布为何呈现如今格局的学科。本文尝试通过对不同地理居群冰清绢蝶翅脉特征的几何形态学计量,推测冰清绢蝶现有分布格局的形成机制,旨为相关领域研究提供参考。

图1冰清绢蝶形态图
1 材料与方法
1.1 实验材料
本文选取冰清绢蝶10个地理居群的标本(每一居群随机抽样20个体),涵括了冰清绢蝶的大部分分布区,具体的采样点信息如表1所示。野外采集的标本立即投入无水乙醇中(无锡市展望化工试剂有限公司)保存,所取标本均来源于安徽师范大学生命科学学院分子系统学与进化研究室样品库内。
1.2标本处理
取出浸泡在无水乙醇中的蝴蝶,用镊子夹取蝶翅的基部,使其完整地脱落,待酒精挥发完全,在蝶翅下方置垫一小方块白纸,便于与黑色翅脉形成反差;其上盖上一载玻片,将蝶翅压平,放入基恩士VHX-5000三维数码显微镜(日本)中观察测量(分辨率2400DPI)。
表1十个地理居群冰清绢蝶采样点信息
序号 | 采样点 | 采集日期 | 抽取标本量 | 地理坐标 |
1 | 江苏南京 | 2011.5.5 | 20 | E 118.83 N 32.06 |
2 | 江苏连云港 | 2011.5.1 | 20 | E 119.40 N 34.71 |
3 | 安徽天堂寨 | 2009.4.29 | 20 | E 115.80 N 31.20 |
4 | 陕西黄柏塬 | 2013.6.8 | 20 | E 107.40 N 33.73 |
5 | 浙江天目山 | 2012.5.3 | 20 | E 119.45 N 30.34 |
6 | 山东泰山 | 2013.5.10 | 20 | E 117.12 N 36.25 |
7 | 甘肃天水 | 2013.6.12 | 20 | E 105.68 N 34.85 |
8 | 安徽琅琊山 | 2013.5.5 | 20 | E 118.29 N 32.28 |
9 | 湖北神农架 | 2014.5.7-10 | 20 | E 110.35 N 31.52 |
10 | 河南嵩山 | 2016.5.5 | 20 | E 113.05 N 34.49 |
1.3图像获取
利用基恩士VHX-5000三维数码显微镜的图像拼接技术对图像进行采集,形成一张完整的蝴蝶全翅图片,在该数码显微镜的测量工具中,利用鼠标多点连线或两点间连线描摹出翅脉的纹路并且勾勒出所需测量的翅脉长度和面积,即可进行测量。前翅共测得13组数据,后翅共测得4组数据,保存拍摄图片以及测量结果生成扩展名为.csv的文件,利用VHX Measurement Data Tabulation Tool软件将数据导出至Excel表格中。
1.4数据分析
在Excel表格中,整理和统计10个居群共200个绢蝶所测得的翅脉长度和面积,并求出均值,对翅脉长度、蝶翅中室面积、全翅面积等性状进行统计分析,利用SPSS 21.0软件分别对各组数据进行初步系统聚类,筛选出聚类结果相同的一些性状,舍弃部分不适合聚类分析的性状,最后对筛选出的性状进行统计和聚类分析。
2结果与分析
2.1十个冰清绢蝶地理居群的翅脉形态学特征平均值
分别取10个地理居群各20个体冰清绢蝶,测量前翅11组长度数据、2组面积数据;测量后翅2组长度数据、2组面积数据(图2)。求得前后翅翅脉形态学特征平均值(见表2,表3),表2为前翅翅脉数据,表3为后翅翅脉数据。根据Excel文档求出P值结果小于0.05,表示各特征值的不同均由不同居群的冰清绢蝶的形态差异引起。
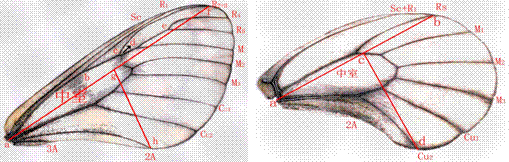
图2冰清绢蝶翅脉交点位置分布图(左.前翅;右.后翅)
从表中可以看出南京、连云港、天堂寨、天目山的冰清绢蝶蝶翅面积较大,各翅脉长度相对较长。利用SPSS 21.0对前后翅共计17组数据进行聚类分析,并一一比对,选出聚类分析较为相似的数据共10组,即为有效性状。
表2十个地理居群冰清绢蝶前翅脉形态学特征平均值
地区 | 中室面积(μm2) | 全翅面积(μm2) | Sc(μm) | ab(μm) | R1(μm) | bc(μm) | de(μm) | R4(μm) | R5(μm) | M1(μm) | Cu2(μm) | 2A(μm) | af(μm) |
江苏南京 | 79097950 | 512958635 | 27969 | 15076 | 15798 | 6356 | 7827 | 9453 | 9604 | 14331 | 15236 | 20807 | 35806 |
江苏连云港 | 75535418 | 461392131 | 26222 | 14556 | 14745 | 5655 | 6619 | 10194 | 10095 | 13431 | 14203 | 19518 | 34329 |
安徽天堂寨 | 80678170 | 502729246 | 27886 | 14809 | 15842 | 6393 | 7158 | 10311 | 10181 | 13903 | 14752 | 19907 | 35862 |
陕西黄柏塬 | 69103127 | 436490081 | 26084 | 13680 | 15076 | 6036 | 6698 | 9385 | 9368 | 12450 | 13760 | 18985 | 33456 |
浙江天目山 | 79277011 | 474916285 | 26743 | 14610 | 15323 | 6178 | 6450 | 9646 | 9774 | 13406 | 14261 | 19157 | 34347 |
山东泰山 | 70204164 | 443461663 | 25097 | 13702 | 14455 | 5985 | 6642 | 9337 | 9411 | 12484 | 13700 | 18307 | 32741 |
甘肃天水 | 71899058 | 449518361 | 26344 | 14018 | 14935 | 5995 | 6655 | 9663 | 9363 | 12504 | 13955 | 18849 | 33713 |
安徽琅琊山 | 68786897 | 441266415 | 25661 | 13786 | 14571 | 5586 | 7167 | 9286 | 9485 | 13396 | 14177 | 18851 | 32985 |
湖北神农架 | 67560787 | 433908146 | 26380 | 14654 | 14581 | 5404 | 6663 | 9338 | 9426 | 13071 | 14184 | 19931 | 33462 |
河南嵩山 | 68035854 | 428151714 | 24217 | 13435 | 14276 | 5763 | 6649 | 9239 | 9511 | 12783 | 13561 | 18434 | 32213 |
表3十个地理居群冰清绢蝶后翅脉形态学特征平均值
地区 | ab(μm) | cd(μm) | 中室面积(μm2) | 全翅面积(μm2) | ||
江苏南京 | 25394 | 16276 | 69996228 | 444529813 | ||
江苏连云港 | 24172 | 14780 | 63192952 | 393275400 | ||
安徽天堂寨 | 25279 | 15223 | 66969326 | 434725131 | ||
陕西黄柏塬 | 23124 | 14567 | 57916839 | 369232093 | ||
浙江天目山 | 24631 | 15340 | 67012411 | 412263624 | ||
山东泰山 | 23192 | 14395 | 56853332 | 374894344 | ||
甘肃天水 | 23594 | 14395 | 59670751 | 377557913 | ||
安徽琅琊山 | 23683 | 14698 | 56607978 | 376142857 | ||
湖北神农架 | 22804 | 14114 | 54779882 | 348904919 | ||
河南嵩山 | 22839 | 14218 | 57192811 | 359625240 |
2.2十个冰清绢蝶地理居群的聚类分析树状图
利用SPSS 21.0再将10组有效性状(前翅分别为ab、R1、R5、M1、Cu2、2A、af、中室面积;后翅分别为ab、中室面积)进行聚类分析,绘制出树状图,从树状图得出结论:冰清绢蝶的演化划分为两支,聚为两大类,一类为天堂寨、南京、天目山、连云港四个地理居群;另一类为琅琊山、泰山、嵩山、天水、黄柏塬、神农架六个地理居群。聚为一类的显示亲缘关系较近。(图3)。
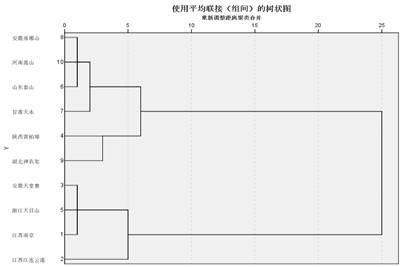
图3冰清绢蝶10个地理居群的聚类分析树状图
2.3冰清绢蝶演化路径推测图
根据2.2聚类树状图绘制成冰清绢蝶演化路径的推测图(图4)。地质学资料显示,大约60百万年以前,印度板块和亚欧板块碰撞导致的造山运动逐渐形成了青藏高原,包括喜马拉雅山脉[11]。同时,相关的分子系统学和分子钟研究推测,距今大约20~30百万年前,绢蝶属物种的祖先在我国西南地区(现今藏东南的横断山脉一带)开始形成[8],同时产生了一些适应高海拔环境的特性,如身翅被毛,耐寒冷、耐低氧等,随着绢蝶属的分化和物种之间的竞争,以及中新世以后,特别是第四纪期间全球气候变化的影响,冰清绢蝶开始向华中、华北、华东地区迁移,形成两支演化路径:一支由喜马拉雅山脉经过唐古拉山、巴颜喀拉山,
到达位于秦岭的天水、黄柏塬、神农架一带,再分为三小支,途经华北平原分别向泰山、嵩山、琅琊山三个方向迁移;另一支由喜马拉雅山脉发源,向东迁移经过秦岭、四川盆地到达天堂寨、南京、天目山等地,最后沿海岸线扩散至连云港一带。成为我国唯一一种分布到长江以南,生活在较低海拔地区的绢蝶,且对生活环境要求极为苛刻[7,12,13]。
3讨论
安徽天堂寨位于大别山区,是南北水系的分水岭,属大别山系,地质基础为前震旦纪的秦岭褶皱带,后期受燕山期造山运动的显著影响;浙江天目山和江苏南京紫金山总体上属于宁镇山脉,是侏罗纪末到白垩纪初期,通过宁镇山脉造山运动而形成的一条向北突出的弧形皱;江苏连云港云台山的地质基础则属于华北古陆构架。这些古地质事件形成的山脉后来都经过了频繁的地质变化过程才最终形成了现在的地质地貌。
甘肃天水属小陇山系,为秦岭的西北缘;陕西黄柏塬地为秦岭腹地;湖北神农架属大巴山系,河南嵩山属伏牛山系,它们都是秦岭造山带的余脉;山东泰山构造基础为古华北地带,后来经漫长而又复杂的演化过程才形成今天的地貌;安徽琅琊山为江淮之间的低山丘陵地带,系大别山系向东延伸而成的余脉。
总之,上述地质过程,包括较晚近的燕山运动时期(在地史上主要属于侏罗纪末到古近纪初这段时期,距今2亿1千万年左右开始到6500万年前为止)相对于绢蝶种群的分化时期而言(据分子钟推测为350万前至30万年前)是一个十分古老的时期,因而,这些地质因素对冰清绢蝶的扩散无关。
综合多方面证据分析,笔者推测,冰清绢蝶不同居群间的扩散路径与其地理分布(相关山脉分布、走向及地理隔离等)之间有明显的相关性,即地质历史上,第四纪开始至今的地质、生物环境和气候变化因素对冰清绢蝶的扩散路径和现代地理分布产生了重要影响[8,11]。譬如,第一进化支(天堂寨、天目山、南京、连云港四个地理居群)各居群的生态环境极为相似,均位于华北平原与长江中下游平原地带,海拔较低,降水丰富,属于亚热带季风气候与温带季风气候交界处;而第二进化支(天水、黄柏塬、神农架、嵩山、泰山五个地理居群)的生态环境类似,海拔相对较高、气温也相对较低,地处内陆,降水相对较少。至于安徽琅琊山的居群,单从地理因素考虑,理论上应属于第一进化支,这与安徽师范大学生命科学学院分子系统学与进化研究室利用分子手段分析的结果基本一致(实验结果暂未发表)。但从形态上看,琅琊山居群确实很特殊(如后翅基部较大范围明显的黑色斑块,体毛为灰白色),且和嵩山或泰山等高纬度地区的居群极为相似。笔者推测这有可能受其寄主植物及采蝶时(2013年4月29日)当年的气候影响,因为不同气候(如阴雨、干旱、季节变化等),寄主植物均影响蝶类个体发生和发育[12,14-16,17]。
笔者查阅了冰清绢蝶寄主植物和气象的相关资料。资料显示,延胡索是琅琊山冰清绢蝶的唯一寄主植物,且大都分布在琅琊山山顶(海拔200-300m),且蝶类对寄主植物专一性较强,虽然它们有一定的迁飞能力,但其分布仍然以寄主为中心[12,13,18,19]。据文献记载,延胡索琅琊山居群的6种主要异喹啉生物碱含量在河南、安徽、浙江、江苏、江西等12个居群中大都高于其它居群含量。尤其是紫堇碱含量最高(0.44%),比含量最低的江西宜兴大滩岭居群(0.024%)高达近20倍,同时延胡索对环境要求很严,对于低洼易积水及水分较少的干旱地段都不适合其生长,生长期仅有3个月,一般分布在海拔200—800米的丘陵山区的半阴坡,系早春植物[20];同时从安徽滁州气象站提供气象资料和中央气象局数据库数据显示,琅琊山地区2013年3、4月降水量均为18.4mm,与近三十年同期均值相比,偏少7成;日照均为240小时,与常年相比偏多4成,整体气候近为春旱,冰清绢蝶与其寄主植物休戚相关。所以,琅琊山居群独特的翅脉形态特征可能与其寄主植物和当地的气候条件等都有密切关系。
4小结
本文通过对10个不同地理居群冰清绢蝶翅脉几何形态学的初步分析显示,冰清绢蝶不同居群间的扩散路径与其地理分布之间有着明显的相关性,其扩散路径及进化有两个分支,聚为两大类,一支是天堂寨、天目山、琅琊山、南京、连云港五个地理居群聚为一类,显示亲缘关系近;一支是天水、黄柏塬、神农架、泰山、嵩山五个地理居群聚为一类,显示亲缘关系近。本研究完全支持安徽师范大学生命科学学院分子系统学与进化研究室利用分子手段分析的结果。研究结果表明,几何形态学分析方法在蝶类的种内分化研究中也是一种十分可行简易的有效手段,对于探讨生物的个体发育、种群分化、系统进化等多方面很有意义和价值。
[参考文献]
[1]蔡小娜,韩旭,沈佐锐,等.基于蛾翅脉特征的夜蛾昆虫数字化分类研究[J].环境昆虫学报, 2016, 38(2):348-353.
[2] GE De-Yan, XIA Lin, LV Xue-Fei,et al. Methods in geometric morphometrics and their applications in ontogenetic and evolutionary biology of animals[J]. Acta Zootaxonomica Sinica, 2012, 37(2):296-304.
[3]蔡小娜,高灵旺,黄大庄,等.利用翅的数学形态特征对蛾类昆虫进行分类鉴定的系统研究Ⅱ——在科级阶元上的应用[J].中国农业大学学报,2013,18(5):96-103.
[4] 潘鹏亮,杨红珍,沈佐锐,等. 翅脉的数学形态特征在蝴蝶分类鉴定中的应用研究[J]. 昆虫分类学报, 2008, 30(2):151-160.
[5] 潘鹏亮,杨红珍,沈佐锐,等. 三种绢蝶翅脉数字化特征的提取及初步分析[J].动物分类学报, 2008, 33(3):566-571.
[6]周尧.中国蝶类志[M].郑州:河南科学技术出版社, 2000:191-193,199,783.
[7]吴琦.冰清绢蝶(上)[J].大自然,1998(3)17-20.
[8]丁亮,张彦周,朱朝东.锯凤蝶类和绢蝶类(麟翅目,凤蝶科)分类地位及谱系关系初探[J].动物分类学报, 2007,32(2):355-362.
[9] Toru Katoh, Anton Chichvarkhin,Takashi Yagi ,et al. Phylogeny and Evolution of Butterflies of the Genus Parnassius;Inferences from Mitochondrial 16S and ND1 Sequences [J]. Zoological Society of Japan, 2005(22):343-351.
[10]吴琦.冰清绢蝶(下)[J].大自然, 1998, (4):19-23.
[11]马铁山,郝改莲,刘殿锋,等.中国凤蝶科昆虫地理分布的聚类分析[J].昆虫知识, 2009,64(4):615-619.
[12] 诸立新,颜劲松,郑和权,等.安徽琅琊山蝶类季节变化的研究[J].滁州师专学报, 2003, 5(4): 95-97.
[13]张起玉,申卫星,李艳,等.泰山蝶类物种多样性及区系分析[J].山东林业科技, 2011, 41(3):
29-32.
[14]郝祥瑜,孙祯,徐震,陶红梅.我国主要蝴蝶地理区域多样性的研究性学习[J]. 中学生物学,2015,31(6):36-38.
[15] 许国权,段海生,刘亦仁. 大别山主峰天堂寨地区蝶类资源及区系组成研究[J].湖北大学学报(自然科学版), 2010,32(3):330-334.
[16]李朝辉,宋东杰,赵清良,等.南京紫金山地区鳞翅目蝶类种类分布及其季节性变化[J].四川动物, 2001, 20(2):76-78.
[17]张如力.甘肃省绢蝶属的种类及区系[J].草业学报, 2005, 14(1):49-52.
[18]诸立新,吴孝兵,欧永跃.天目山北坡蝶类资源和区系[J].安徽师范大学学报(自然科学版), 2006, 29(3):266-271.
[19]涂清芳.琅琊山风景区野生观赏植物资源调查及评价[D].南京:南京林业大学, 2012.
[20]许翔鸿,余国奠,王峥涛.野生延胡索种质资源现状及其质量评价[J]. 2004, 29(5):399-401.
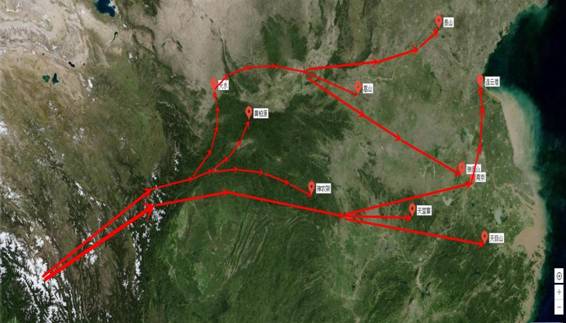 图4
图4
